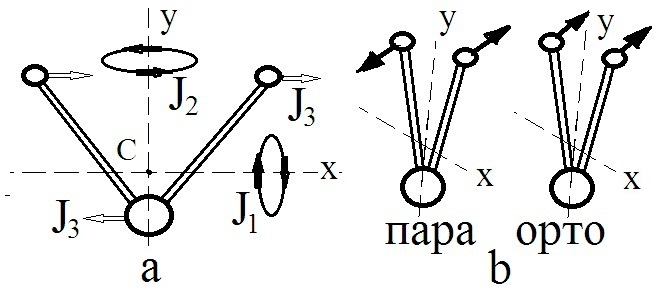
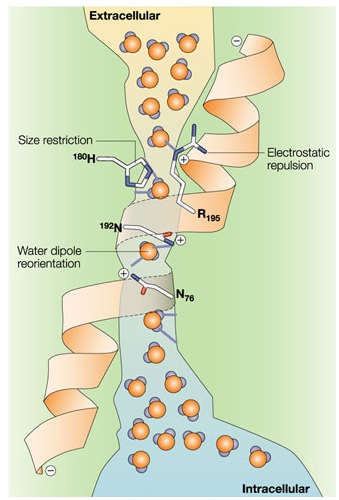
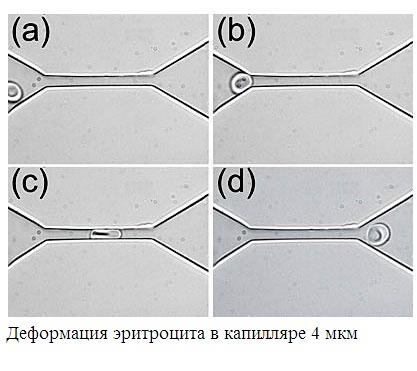
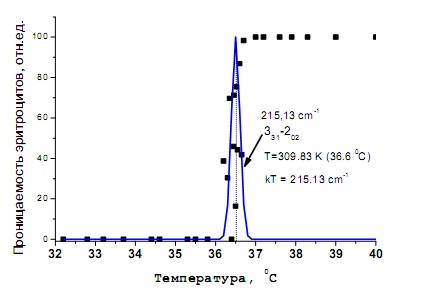
ХИРАЛЬНОСТЬ И КВАНТОВЫЕ ЭФФЕКТЫ КАК ФАКТОРЫ МОРФОГЕНЕЗА
А.С. Холманский
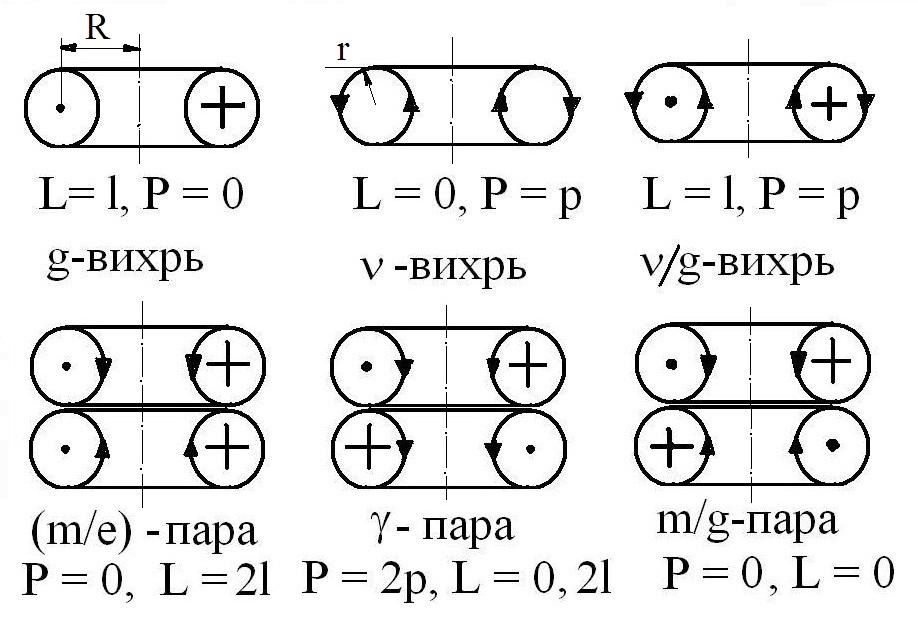
- формализм фрактально-резонансных изоэнергетических обратимых трансформаций или флуктуаций ЭФ эфира:
(1)

декстрана и скипидара от температуры и даты [19]. Снижение
раствора декстрана при жаркой погоде в июле августе в Подмосковье обусловлено с распадом молекулярных комплексов, имеющих более высокое значение удельного вращения, по сравнению с олигомерами декстрана [10]. Энергия активации реакции образования комплексов для различных сахаров меняется в пределах 0,1 - 0,6 кДж/моль и изменение температуры на 10оС уже дает ощутимый эффект [10].
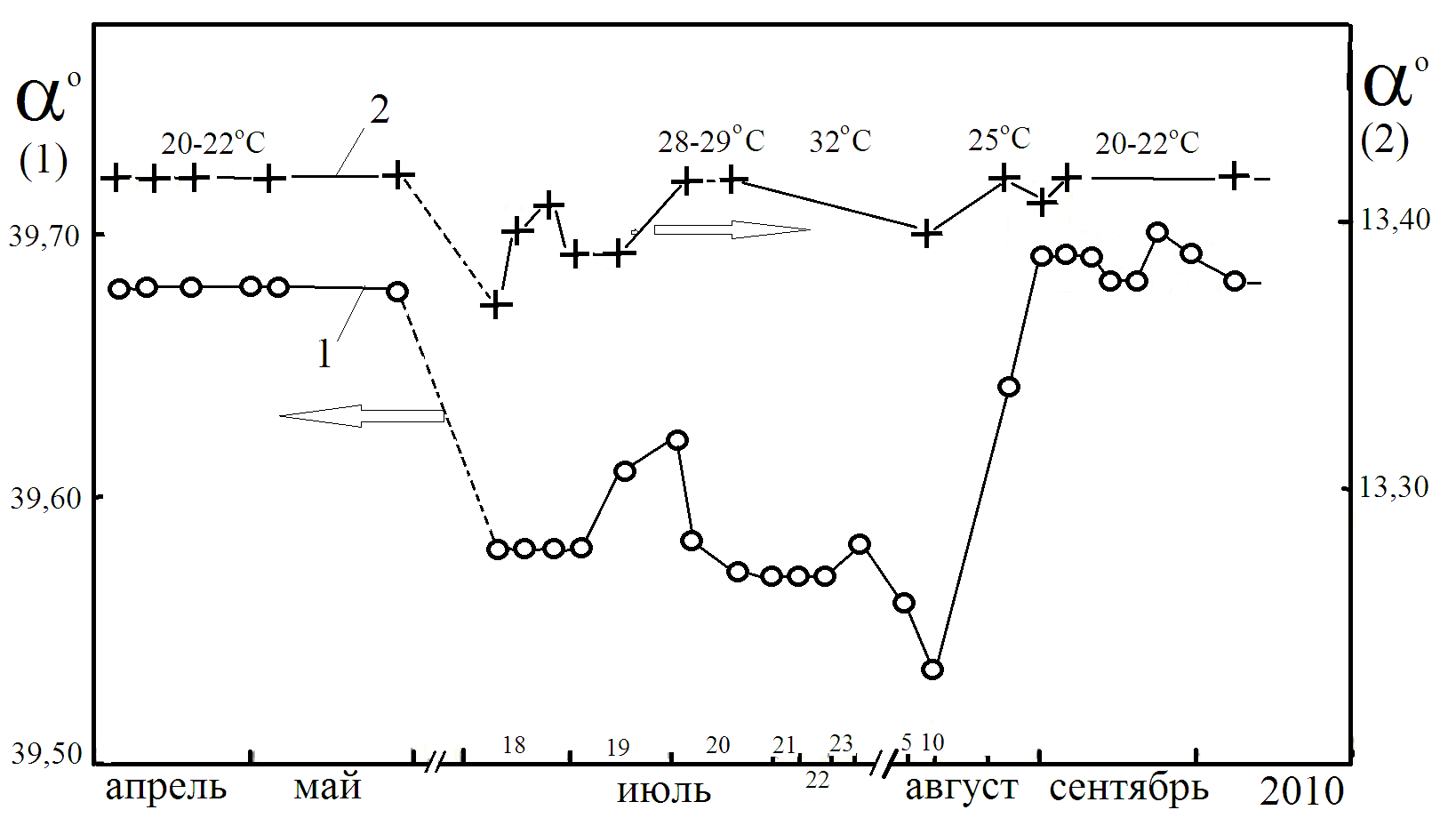
раствора декстрана и скипидара, например, 21 июля и 9 августа могут быть обусловлены изменением магнитной обстановки на Земле вследствие повышения активности Солнца - в эти дни существенно возрастала суммарная площадь солнечных пятен и возрастала интенсивность радиоизлучения на длине волны 10,7 см (по данным ИЗМИРАН).