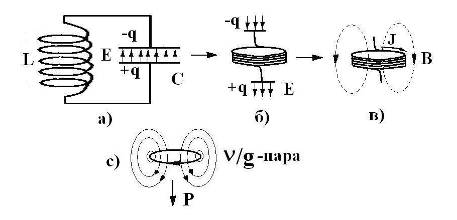
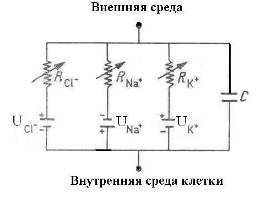
Для объяснения электрических свойств мембраны привлекают схему эквивалентного контура, в которой проводящие каналы для различных ионов моделируют источником ЭДС и омическим сопротивлением (R), а изоляционные свойства мембраны представляют емкостью (Рис 2).
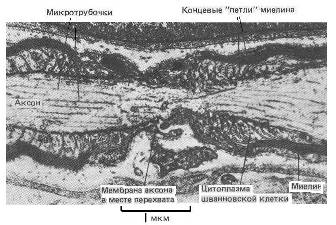
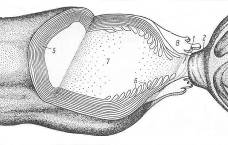
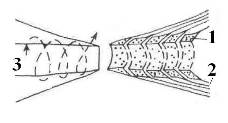
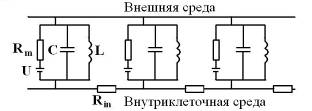
Рис 8. Модифицированная электрическая схема мембраны нерва. Rm , U - ионный канал; С - емкость мембраны; L - индуктивность глиальных миелиновых спиралей мембраны; Rin - сопротивление аксоплазмы.
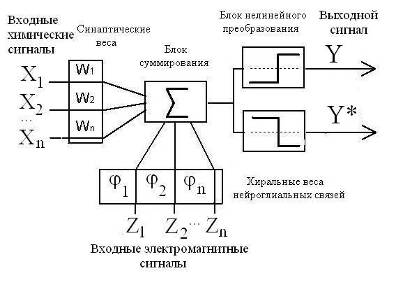
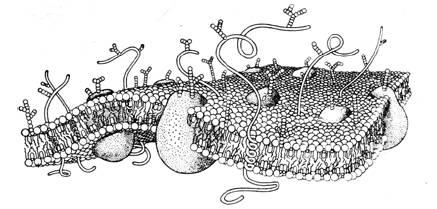
В силу этого следует предполагать существенное влияние эпитаксиального эффекта на процессы, регулирующие межнейронные и нейроглиальные взаимодействия. Известно, например, что в химических реакциях, протекающих в оптически активной среде или на поверхности кварца, возрастает выход хиральных продуктов. Увеличению эпитаксиального эффекта мембран и стенок различных органов, помимо посредничества воды, очевидно, способствуют связанные или адсорбированные поверхностью полипептидные и полисахаридные цепочки, а также микроворсинки (Рис 12) [15, 17]. Эпитаксиальный эффект и присутствие хиральных сахаров сказывается на кинетике обратимой адсорбции ионов и нейромедиаторов на поверхностях мембран нейронов как в перехватах Ранвье, так и в синапсах [14].
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
С6Н12О6 + 6 О2 ? 6 СО2 + 6 Н2О + 2800 (кДж/моль). (12)